






კონცეფცია, რომელიც დღეს ცნობილია, როგორც PRP, პირველად გამოჩნდა ჰემატოლოგიის სფეროში 1970-იან წლებში.ჰემატოლოგებმა შექმნეს ტერმინი PRP ათწლეულების წინ, რათა აღწერონ პლაზმა, რომელიც მიღებულია პერიფერიულ სისხლში ბაზალური მნიშვნელობების ზემოთ თრომბოციტების რაოდენობის მიხედვით.ათ წელზე მეტი ხნის შემდეგ, PRP გამოიყენებოდა ყბა-სახის ქირურგიაში, როგორც თრომბოციტებით მდიდარი ფიბრინის (PRF) ფორმა.ამ PRP წარმოებულში ფიბრინის შემცველობა დიდი მნიშვნელობა აქვს მისი წებოვანი და ჰომეოსტატიკური თვისებების გამო, ხოლო PRP-ს აქვს მდგრადი ანთების საწინააღმდეგო თვისებები და ასტიმულირებს უჯრედების პროლიფერაციას.საბოლოოდ, დაახლოებით 1990-იან წლებში, PRP პოპულარული გახდა და საბოლოოდ, ტექნოლოგია სხვა სამედიცინო სფეროებშიც გადავიდა.მას შემდეგ ეს პოზიტიური ბიოლოგია ფართოდ იქნა შესწავლილი და გამოყენებული პროფესიონალ სპორტსმენებში კუნთოვანი სისტემის სხვადასხვა დაზიანებების სამკურნალოდ, რაც შემდგომში ხელს უწყობს მის ფართო მედია ყურადღებას.გარდა იმისა, რომ ეფექტურია ორთოპედიასა და სპორტულ მედიცინაში, PRP გამოიყენება ოფთალმოლოგიაში, გინეკოლოგიაში, უროლოგიასა და კარდიოლოგიაში, პედიატრიაში და პლასტიკურ ქირურგიაში.ბოლო წლებში PRP-ს ასევე აფასებენ დერმატოლოგები კანის წყლულების სამკურნალოდ, ნაწიბურების რეგენერაცია, კანის გაახალგაზრდავება და თმის ცვენაც კი.
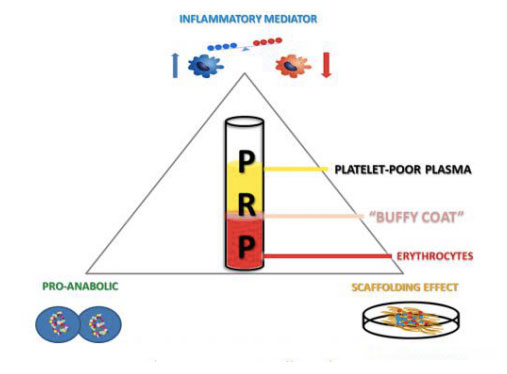
იმის გათვალისწინებით, რომ ცნობილია, რომ PRP პირდაპირ მანიპულირებს შეხორცებით და ანთებითი პროცესებით, სამკურნალო კასკადი უნდა იყოს დანერგილი, როგორც მითითება.შეხორცების პროცესი იყოფა შემდეგ ოთხ ეტაპად: ჰემოსტაზი;ანთება;უჯრედული და მატრიქსის პროლიფერაცია და ბოლოს ჭრილობის რემოდელირება.
1. ქსოვილის შეხორცება
გააქტიურებულია ქსოვილის შეხორცების კასკადი, პროცესი, რომელიც იწვევს თრომბოციტების აგრეგაციას, თრომბის წარმოქმნას და დროებითი უჯრედგარე მატრიქსის განვითარებას (ECM. თრომბოციტები შემდეგ ეკვრის გამოვლენილ კოლაგენს და ECM ცილებს, რაც იწვევს α-გრანულების არსებობას გათავისუფლებაში. ბიოაქტიური მოლეკულები. თრომბოციტები შეიცავს მრავალფეროვან ბიოაქტიურ მოლეკულებს, მათ შორის ზრდის ფაქტორებს, ქიმიოკინებს და ციტოკინებს, ასევე ანთების საწინააღმდეგო შუამავლებს, როგორიცაა პროსტაგლანდინები, პროსტატის ციკლინი, ჰისტამინი, თრომბოქსანი, სეროტონინი და ბრადიკინინი.
შეხორცების პროცესის საბოლოო ეტაპი დამოკიდებულია ჭრილობის რემოდელირებაზე.ქსოვილის რემოდელირება მკაცრად რეგულირდება ანაბოლურ და კატაბოლურ პასუხებს შორის ბალანსის დასამყარებლად.ამ ფაზის განმავლობაში, თრომბოციტებიდან მიღებული ზრდის ფაქტორი (PDGF), ტრანსფორმაციული ზრდის ფაქტორი (TGF-β) და ფიბრონექტინი ასტიმულირებს ფიბრობლასტების პროლიფერაციას და მიგრაციას, ასევე ECM კომპონენტების სინთეზს.თუმცა, ჭრილობის მომწიფების დრო დიდწილად არის დამოკიდებული ჭრილობის სიმძიმეზე, ინდივიდუალურ მახასიათებლებზე და დაზიანებული ქსოვილის სპეციფიკურ შეხორცების შესაძლებლობებზე და ზოგიერთმა პათოფიზიოლოგიურმა და მეტაბოლურმა ფაქტორებმა შეიძლება გავლენა მოახდინონ შეხორცების პროცესზე, როგორიცაა ქსოვილის იშემია, ჰიპოქსია, ინფექცია. , ზრდის ფაქტორების დისბალანსი და მეტაბოლურ სინდრომთან დაკავშირებული დაავადებებიც კი.
ანთების საწინააღმდეგო მიკროგარემო, რომელიც ხელს უშლის შეხორცების პროცესს.საქმეების გასართულებლად, ასევე არსებობს პროტეაზას მაღალი აქტივობა, რომელიც აფერხებს ზრდის ფაქტორის (GF) ბუნებრივ მოქმედებას.მიტოგენური, ანგიოგენური და ქიმიოტაქსიური თვისებების გარდა, PRP ასევე არის მრავალი ზრდის ფაქტორის მდიდარი წყარო, ბიომოლეკულები, რომლებსაც შეუძლიათ წინააღმდეგობა გაუწიონ მავნე ეფექტებს ანთებით ქსოვილებში გამწვავებული ანთების კონტროლით და ანაბოლური სტიმულის დამყარებით.ამ თვისებების გათვალისწინებით, მკვლევარებმა შეიძლება იპოვონ დიდი პოტენციალი სხვადასხვა რთული დაზიანებების მკურნალობაში.
2. ციტოკინი
ციტოკინები PRP-ში მთავარ როლს ასრულებენ ქსოვილების აღდგენის პროცესების მანიპულირებაში და ანთებითი დაზიანების რეგულირებაში.ანთების საწინააღმდეგო ციტოკინები არის ბიოქიმიური მოლეკულების ფართო სპექტრი, რომლებიც შუამავლობენ ანთების პრო-ანთებით ციტოკინის პასუხებს, ძირითადად გამოწვეული გააქტიურებული მაკროფაგებით.ანთების საწინააღმდეგო ციტოკინები ურთიერთქმედებენ სპეციფიკურ ციტოკინის ინჰიბიტორებთან და ხსნად ციტოკინის რეცეპტორებთან ანთების მოდულირებისთვის.ინტერლეუკინის (IL)-1 რეცეპტორის ანტაგონისტები, IL-4, IL-10, IL-11 და IL-13 კლასიფიცირდება, როგორც ძირითადი ანთების საწინააღმდეგო ციტოკინები.ჭრილობის ტიპებიდან გამომდინარე, ზოგიერთ ციტოკინს, როგორიცაა ინტერფერონი, ლეიკემიის ინჰიბიტორული ფაქტორი, TGF-β და IL-6, შეუძლია გამოავლინოს პრო- ან ანთების საწინააღმდეგო ეფექტი.TNF-α, IL1 და IL-18 აქვთ ციტოკინის გარკვეული რეცეპტორები, რომლებიც შეიძლება დათრგუნონ სხვა ცილების პრო-ანთებით ეფექტებს [37].IL-10 არის ერთ-ერთი ყველაზე ძლიერი ანთების საწინააღმდეგო ციტოკინი, მას შეუძლია შეამციროს ანთების საწინააღმდეგო ციტოკინები, როგორიცაა IL-1, IL-6 და TNF-α, და არეგულირებს ანთების საწინააღმდეგო ციტოკინებს.ეს კონტრრეგულატორული მექანიზმები კრიტიკულ როლს ასრულებენ პრო-ანთებითი ციტოკინების წარმოებასა და ფუნქციონირებაში.გარდა ამისა, ზოგიერთმა ციტოკინმა შეიძლება გამოიწვიოს სპეციფიკური სასიგნალო რეაქციები, რომლებიც ასტიმულირებენ ფიბრობლასტებს, რომლებიც გადამწყვეტია ქსოვილის აღდგენისთვის.ანთებითი ციტოკინები TGFβ1, IL-1β, IL-6, IL-13 და IL-33 ასტიმულირებენ ფიბრობლასტებს, რათა დიფერენცირდნენ მიოფიბრობლასტებად და გააუმჯობესონ ECM [38].თავის მხრივ, ფიბრობლასტები გამოყოფენ ციტოკინებს TGF-β, IL-1β, IL-33, CXC და CC ქიმიოკინებს, რომლებიც ხელს უწყობენ ანთების საწინააღმდეგო რეაქციებს იმუნური უჯრედების გააქტიურებით და რეკრუტირებით, როგორიცაა მაკროფაგები.ამ ანთებით უჯრედებს აქვთ მრავალი როლი ჭრილობის ადგილზე, უპირველეს ყოვლისა, ჭრილობის გაწმენდის ხელშეწყობით - ასევე ქიმიოკინების, მეტაბოლიტების და ზრდის ფაქტორების ბიოსინთეზის გზით, რაც აუცილებელია ახალი ქსოვილის რემოდელირებისთვის.ამრიგად, PRP-ში არსებული ციტოკინები მნიშვნელოვან როლს ასრულებენ უჯრედის ტიპის შუამავლობით გამოწვეული იმუნური რეაქციების სტიმულირებაში, რაც ხელს უწყობს ანთებითი ფაზის გადაწყვეტას.ფაქტობრივად, ზოგიერთმა მკვლევარმა ამ პროცესს უწოდა "რეგენერაციული ანთება", რაც ვარაუდობს, რომ ანთებითი ფაზა, მიუხედავად პაციენტის შფოთვისა, არის კრიტიკული ნაბიჯი, რომელიც აუცილებელია ქსოვილის აღდგენის პროცესისთვის წარმატებული დასკვნის მისაღწევად, ეპიგენეტიკური მექანიზმების გათვალისწინებით, რომლითაც ანთებითი სიგნალები ხელს უწყობს უჯრედულ მუშაობას. პლასტიურობა.
3. ფიბრინი
თრომბოციტები ატარებენ ფიბრინოლიზურ სისტემასთან დაკავშირებულ რამდენიმე ფაქტორს, რამაც შეიძლება მოახდინოს ფიბრინოლიზური პასუხის რეგულაცია ან დაქვეითება.ჰემატოლოგიური კომპონენტებისა და თრომბოციტების ფუნქციის დროებითი ურთიერთობა და შედარებითი წვლილი თრომბოციტების დეგრადაციაში რჩება საკითხად, რომელიც იმსახურებს ფართო განხილვას საზოგადოებაში.ლიტერატურაში წარმოდგენილია მრავალი კვლევა, რომელიც ფოკუსირებულია მხოლოდ თრომბოციტებზე, რომლებიც ცნობილია სამკურნალო პროცესზე გავლენის მოხდენის უნარით.მრავალი გამორჩეული კვლევის მიუხედავად, სხვა ჰემატოლოგიური კომპონენტები, როგორიცაა კოაგულაციის ფაქტორები და ფიბრინოლიზური სისტემა, ასევე მნიშვნელოვანი წვლილი შეიტანეს ჭრილობის ეფექტურ აღდგენაში.განმარტებით, ფიბრინოლიზი არის რთული ბიოლოგიური პროცესი, რომელიც ეყრდნობა გარკვეული ფერმენტების გააქტიურებას ფიბრინის დეგრადაციის გასაადვილებლად.ფიბრინოლიზური პასუხი შემოთავაზებულია სხვა ავტორების მიერ, რომ ფიბრინის დეგრადაციის პროდუქტები (fdp) შეიძლება რეალურად იყოს მოლეკულური აგენტები, რომლებიც პასუხისმგებელნი არიან ქსოვილის აღდგენის სტიმულირებაზე, მნიშვნელოვანი ბიოლოგიური მოვლენების თანმიმდევრობაზე ფიბრინის დეპონირებამდე და ანგიოგენეზიდან ამოღებაზე, რაც აუცილებელია ჭრილობის შეხორცებისთვის.თრომბის წარმოქმნა ტრავმის შემდეგ მოქმედებს როგორც დამცავი ფენა, რომელიც იცავს ქსოვილს სისხლის დაკარგვისგან, მიკრობული აგენტების შეჭრისგან და ასევე უზრუნველყოფს დროებით მატრიქსს, რომლის მეშვეობითაც უჯრედებს შეუძლიათ მიგრაცია შეკეთების დროს.შედედება განპირობებულია ფიბრინოგენის დაშლით სერინის პროტეაზებით და თრომბოციტების აგრეგატი ჯვარედინი ფიბრინის ბოჭკოვანი ქსელით.ეს რეაქცია იწყებს ფიბრინის მონომერების პოლიმერიზაციას, რაც მთავარი მოვლენაა სისხლის შედედების წარმოქმნაში.თრომბებს შეუძლიათ აგრეთვე იმოქმედონ ციტოკინებისა და ზრდის ფაქტორების რეზერვუარებად, რომლებიც გამოიყოფა გააქტიურებული თრომბოციტების დეგრანულაციისას.ფიბრინოლიზური სისტემა მჭიდროდ რეგულირდება პლაზმინის მიერ და თამაშობს მთავარ როლს უჯრედების მიგრაციის ხელშეწყობაში, ზრდის ფაქტორის ბიოშეღწევადობასა და სხვა პროტეაზას სისტემების რეგულირებაში, რომლებიც მონაწილეობენ ქსოვილების ანთებასა და რეგენერაციაში.ფიბრინოლიზის ძირითადი კომპონენტები, როგორიცაა უროკინაზას პლაზმინოგენის აქტივატორი რეცეპტორი (uPAR) და პლაზმინოგენის აქტივატორის ინჰიბიტორი-1 (PAI-1), ცნობილია, რომ გამოხატულია მეზენქიმურ ღეროვან უჯრედებში (MSCs), სპეციალიზებული ტიპის უჯრედები, რომლებიც აუცილებელია ჭრილობის წარმატებული შეხორცებისთვის.
4. უჯრედების მიგრაცია
პლაზმინოგენის გააქტიურება uPA-uPAR ასოციაციის მეშვეობით არის პროცესი, რომელიც ხელს უწყობს ანთებითი უჯრედების მიგრაციას, რადგან ის აძლიერებს უჯრედგარე პროტეოლიზს.იმის გამო, რომ uPAR-ს არ გააჩნია ტრანსმემბრანული და უჯრედშიდა დომენები, ცილას ესაჭიროება კორეცეპტორები, როგორიცაა ინტეგრინები და ვიტრეინები უჯრედების მიგრაციის დასარეგულირებლად.გარდა ამისა, uPA-uPAR-ის შეკავშირებამ გამოიწვია uPAR-ის აფინურობის გაზრდა მინისებრი კონნექსინებისა და ინტეგრინების მიმართ, რაც ხელს უწყობს უჯრედების ადჰეზიას.პლაზმინოგენის აქტივატორის ინჰიბიტორი-1 (PAI-1) თავის მხრივ წყვეტს უჯრედებს, ანადგურებს უპარ-ვიტრეინსა და ინტეგრინს- როდესაც ის უკავშირდება uPA-უპარ-ინტეგრინის კომპლექსის uPA-ს უჯრედის ზედაპირზე. შუშის ვოქსელების ურთიერთქმედება.
რეგენერაციული მედიცინის კონტექსტში, მეზენქიმული ღეროვანი უჯრედები მობილიზებულია ძვლის ტვინიდან ორგანოს მძიმე დაზიანების კონტექსტში და, ამრიგად, შეიძლება აღმოჩნდეს მრავალჯერადი მოტეხილობის მქონე პაციენტების მიმოქცევაში.თუმცა, გარკვეულ გარემოებებში, როგორიცაა თირკმელების ბოლო სტადიის უკმარისობა, ღვიძლის ბოლო სტადიის უკმარისობა ან გულის ტრანსპლანტაციის შემდეგ უარყოფის დაწყებისას, ეს უჯრედები შეიძლება არ იყოს გამოვლენილი სისხლში [66].საინტერესოა, რომ ეს ადამიანის ძვლის ტვინიდან მიღებული მეზენქიმული (სტრომული) წინამორბედი უჯრედები არ შეიძლება გამოვლინდეს ჯანმრთელი ადამიანების სისხლში [67].ასევე ადრე იყო შემოთავაზებული uPAR-ის როლი ძვლის ტვინის მეზენქიმული ღეროვანი უჯრედების მობილიზაციაში, ისევე როგორც ჰემატოპოეზური ღეროვანი უჯრედების (HSC) მობილიზაციისას.ვარაბანენი და სხვ.შედეგებმა აჩვენა, რომ გრანულოციტების კოლონიის მასტიმულირებელი ფაქტორის გამოყენებამ uPAR-დეფიციტურ თაგვებში გამოიწვია MSC-ების უკმარისობა, რაც კვლავ აძლიერებს ფიბრინოლიზური სისტემის დამხმარე როლს უჯრედების მიგრაციაში.შემდგომმა კვლევებმა ასევე აჩვენა, რომ გლიკოზილფოსფატიდილინოზიტოლით დამაგრებული uPA რეცეპტორები არეგულირებს ადჰეზიას, მიგრაციას, პროლიფერაციას და დიფერენციაციას გარკვეული უჯრედშორისი სასიგნალო გზების გააქტიურებით, შემდეგნაირად: ფოსფატიდილინოზიტოლის პრო-გადარჩენის გზა 4,5-ბისფოსფაზა 1/2-კსA და ERK1/2-3. და ადჰეზიური კინაზა (FAK).
MSC-ებმა აჩვენეს შემდგომი მნიშვნელობა ჭრილობის შეხორცების კონტექსტში.მაგალითად, პლაზმინოგენის დეფიციტის მქონე თაგვებმა აჩვენეს სერიოზული შეფერხება ჭრილობების შეხორცების მოვლენებში, რაც მიუთითებს იმაზე, რომ პლაზმინი კრიტიკულად მონაწილეობს ამ პროცესში.ადამიანებში პლაზმინის დაკარგვამ ასევე შეიძლება გამოიწვიოს ჭრილობის შეხორცების გართულებები.სისხლის ნაკადის დარღვევამ შეიძლება მნიშვნელოვნად შეაფერხოს ქსოვილების რეგენერაცია, რაც განმარტავს, თუ რატომ არის ეს რეგენერაციული პროცესები უფრო რთული დიაბეტიან პაციენტებში.
5. მონოციტები და რეგენერაციის სისტემები
ლიტერატურის მიხედვით, ბევრია საუბარი მონოციტების როლზე ჭრილობების შეხორცებაში.მაკროფაგები ძირითადად წარმოიქმნება სისხლის მონოციტებიდან და მნიშვნელოვან როლს ასრულებენ რეგენერაციულ მედიცინაში [81].ვინაიდან ნეიტროფილები გამოყოფენ IL-4, IL-1, IL-6 და TNF-[alpha], ეს უჯრედები, როგორც წესი, შეაღწევენ ჭრილობის ადგილზე დაზიანების შემდეგ დაახლოებით 24-48 საათის შემდეგ.თრომბოციტები ათავისუფლებენ თრომბინს და თრომბოციტების ფაქტორ 4-ს (PF4), ორ ქიმიოკინს, რომლებიც ხელს უწყობენ მონოციტების რეკრუტირებას და მათ დიფერენციაციას მაკროფაგებად და დენდრიტულ უჯრედებად.მაკროფაგების თვალსაჩინო მახასიათებელია მათი პლასტიურობა, ე.ი. მათი უნარი გადაცვალონ ფენოტიპები და ტრანსდიფერენციონ სხვა ტიპის უჯრედებად, როგორიცაა ენდოთელური უჯრედები, რომლებიც შემდგომში აჩვენებენ განსხვავებულ ფუნქციებს სხვადასხვა ბიოქიმიური სტიმულის საპასუხოდ ჭრილობის მიკროგარემოში.ანთებითი უჯრედები გამოხატავენ ორ ძირითად ფენოტიპს, M1 ან M2, რაც დამოკიდებულია ადგილობრივ მოლეკულურ სიგნალზე, რომელიც სტიმულის წყაროა.M1 მაკროფაგები გამოწვეულია მიკრობული აგენტებით და, შესაბამისად, აქვთ უფრო ანთების საწინააღმდეგო ეფექტი.ამის საპირისპიროდ, M2 მაკროფაგები, როგორც წესი, წარმოიქმნება ტიპი 2 პასუხით და აქვთ ანთების საწინააღმდეგო თვისებები, რომლებიც, როგორც წესი, ხასიათდება IL-4, IL-5, IL-9 და IL-13 მატებით.ის ასევე მონაწილეობს ქსოვილების აღდგენაში ზრდის ფაქტორების წარმოქმნით.M1-დან M2 იზოფორმებზე გადასვლა ძირითადად განპირობებულია ჭრილობის შეხორცების შემდგომ ეტაპებზე, სადაც M1 მაკროფაგები იწვევენ ნეიტროფილების აპოპტოზს და იწყებენ ამ უჯრედების კლირენსს.ნეიტროფილების მიერ ფაგოციტოზი ააქტიურებს მოვლენათა ჯაჭვს, რომლის დროსაც ციტოკინის წარმოება გამორთულია, მაკროფაგების პოლარიზაცია და TGF-β1-ის გამოთავისუფლება.ეს ზრდის ფაქტორი არის მიოფიბრობლასტების დიფერენციაციისა და ჭრილობის შეკუმშვის ძირითადი მარეგულირებელი, რაც იძლევა ანთების მოგვარების საშუალებას და პროლიფერაციული ფაზის დაწყებას სამკურნალო კასკადში [57].კიდევ ერთი უაღრესად დაკავშირებული ცილა, რომელიც მონაწილეობს უჯრედულ პროცესებში არის სერინი (SG).დადგინდა, რომ ეს ჰემატოპოეტური უჯრედების მიერ გამოყოფილი გრანულანი აუცილებელია სეკრეციული ცილების შესანახად სპეციფიკურ იმუნურ უჯრედებში, როგორიცაა მასტ უჯრედები, ნეიტროფილები და ციტოტოქსიური T ლიმფოციტები.მიუხედავად იმისა, რომ ბევრი არაჰემატოპოეზური უჯრედი ასევე ასინთეზირებს სეროტონინს, ყველა ანთებითი უჯრედი გამოიმუშავებს ამ ცილის დიდ რაოდენობას და ინახავს მას გრანულებში სხვა ანთებით შუამავლებთან, მათ შორის პროტეაზებთან, ციტოკინებთან, ქიმიოკინებთან და ზრდის ფაქტორებთან შემდგომი ურთიერთქმედებისთვის.უარყოფითად დამუხტული გლიკოზამინოგლიკანის (GAG) ჯაჭვები SG-ში, როგორც ჩანს, კრიტიკულია სეკრეტორული გრანულების ჰომეოსტაზისთვის, რადგან მათ შეუძლიათ დაკავშირება და ხელი შეუწყონ არსებითად დამუხტული გრანულების კომპონენტების შენახვას უჯრედის, პროტეინის და GAG ჯაჭვის სპეციფიკური წესით.რაც შეეხება მათ ჩართულობას PRP-ში, ვულფმა და კოლეგებმა ადრე აჩვენეს, რომ SG-ს დეფიციტი მტკიცედ არის დაკავშირებული თრომბოციტების მორფოლოგიასთან შეცვლილ ცვლილებებთან;თრომბოციტების მე-4 ფაქტორის, ბეტა-თრომგლობულინის და PDGF შენახვის დეფექტები თრომბოციტებში;თრომბოციტების ცუდი აგრეგაცია და სეკრეცია in vitro და თრომბოზი in vivo ქმნის დეფექტებს.ამრიგად, მკვლევარებმა დაასკვნეს, რომ ეს პროტეოგლიკანი, როგორც ჩანს, თრომბოზის მთავარი რეგულატორია.
თრომბოციტებით მდიდარი პროდუქტების მიღება შესაძლებელია ინდივიდის მთლიანი სისხლის შეგროვებით და ცენტრიფუგირებით, ნარევის გამოყოფით სხვადასხვა ფენებად, რომლებიც შეიცავს პლაზმას, თრომბოციტებს, ლეიკოციტებს და ლეიკოციტებს.როდესაც თრომბოციტების კონცენტრაცია ბაზალურ მნიშვნელობებზე მაღალია, ძვლებისა და რბილი ქსოვილების ზრდა შეიძლება დაჩქარდეს მინიმალური გვერდითი ეფექტებით.აუტოლოგიური PRP პროდუქტების გამოყენება შედარებით ახალი ბიოტექნოლოგიაა, რომელიც აგრძელებს პერსპექტიულ შედეგებს სხვადასხვა ქსოვილის დაზიანებების სტიმულირებასა და გაძლიერებულ შეხორცებაში.ამ ალტერნატიული თერაპიული მიდგომის ეფექტურობა შეიძლება მიეკუთვნებოდეს ზრდის ფაქტორების და ცილების ფართო სპექტრის აქტუალურ მიწოდებას, ჭრილობების შეხორცებისა და ქსოვილების აღდგენის ფიზიოლოგიური პროცესების იმიტაციას და მხარდაჭერას.გარდა ამისა, ფიბრინოლიზურ სისტემას აშკარად აქვს მნიშვნელოვანი გავლენა ქსოვილის მთლიან აღდგენაზე.გარდა მისი უნარის შეცვალოს ანთებითი უჯრედების და მეზენქიმული ღეროვანი უჯრედების უჯრედული რეკრუტირება, ის არეგულირებს პროტეოლიზურ აქტივობას ჭრილობების შეხორცების ადგილებში და მეზოდერმული ქსოვილების რეგენერაციის დროს, მათ შორის ძვლის, ხრტილისა და კუნთების ჩათვლით, და ამიტომ არის კუნთოვანი მედიცინის კომპონენტი.
შეხორცების დაჩქარება არის ძალიან მოთხოვნადი მიზანი სამედიცინო სფეროში მრავალი პროფესიონალის მიერ და PRP წარმოადგენს დადებით ბიოლოგიურ ინსტრუმენტს, რომელიც აგრძელებს პერსპექტიულ განვითარებას სტიმულირებასა და რეგენერაციული მოვლენების კარგად კოორდინირებულ ტანდემში.თუმცა, ვინაიდან ეს თერაპიული ინსტრუმენტი რჩება კომპლექსური, განსაკუთრებით იმის გამო, რომ ის ათავისუფლებს უამრავ ბიოაქტიურ ფაქტორს და მათ სხვადასხვა ურთიერთქმედების მექანიზმებს და სასიგნალო ეფექტებს, საჭიროა შემდგომი კვლევები.
(ამ სტატიის შინაარსი ხელახლა იბეჭდება და ჩვენ არ ვაძლევთ რაიმე გამოხატულ ან ნაგულისხმევ გარანტიას ამ სტატიაში მოცემული შინაარსის სიზუსტის, სანდოობის ან სისრულისთვის და არ ვართ პასუხისმგებელი ამ სტატიის მოსაზრებებზე, გთხოვთ, გაიგოთ).
გამოქვეყნების დრო: ივლის-19-2022